



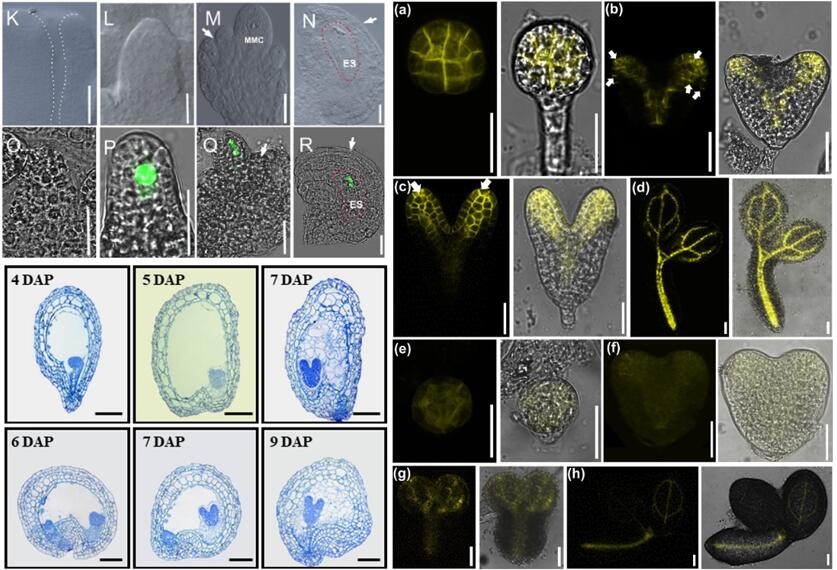
Ovule is the precursor of seed. The number and development of ovule determine the quantity and quality of seed. The development and pattern formation of embryo determine the growth and development after germination. Our group focuses on the regulatory mechanism of the initiation of ovule primordia, the differentiation and development of female gametophyte, and the development and pattern formation of embryo.

Tonoplast proton pumps establish the proton gradient that powers molecular traffic across tonoplast thereby facilitating turgor regulation and nutrient homeostasis. They also affect vacuole morphology and distribution. But how the vacuole and the tonoplast proton pumps influence plant growth and development remains unclear. Our group investigate how the vacuole morphology and distribution and tonoplast proton pumps regulate the d differentiation of germ cells and related reproductive development.
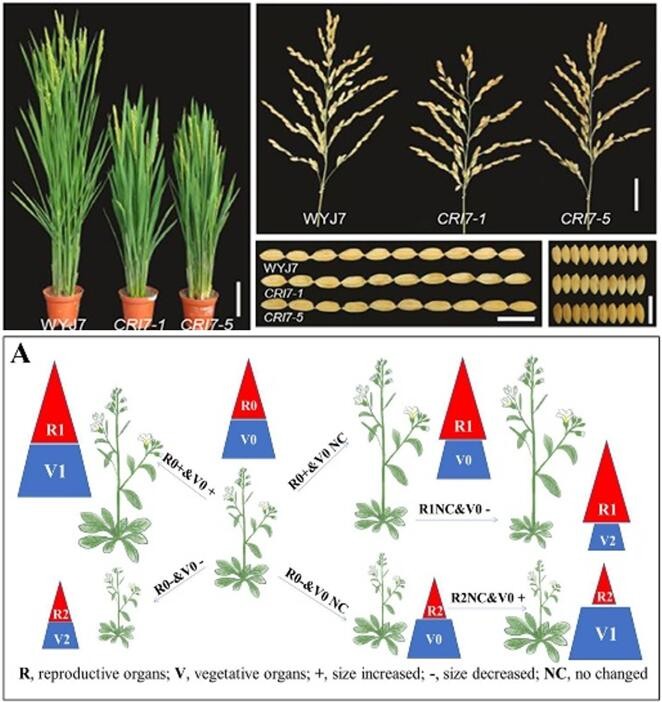
Brassinosteroid (BR) plays crucial roles in modulating plant architecture and seed yield. But BR cannot be applied in agriculture production directly because BR regulated multiple processes and could not be transcriptional regulated in different tissues. The rice mutant with reduced BR signal are dwarf with erect leaves (the ideal architecture for dense planting), but also leads to decreased reproductivity and grain yield. Enhanced BR signaling contributes to increased grain yield, but the increased plant height and leaf inclination induces lodging and reduces planting density. Our group development two solutions, one is identifying new regulators which can mediate parts of BR-regulated rice growth, the other is using tissue specific promoters to derive modified coding sequences to modulate BR signaling directly.
Vacuolar H+-ATPase and BZR1 form a feedback loop to regulate the homeostasis of BR signaling in Arabidopsis. Jiang, Y. T., Yang, L. H., Zheng, J. X., Geng, X. C., Bai, Y. X., Wang, Y. C., ... & Lin, W. H*. (2023). Molecular Plant, 16(12), 1976-1989.
RLI2 regulates Arabidopsis female gametophyte and embryo development by facilitating the assembly of the translational machinery. Yu, S. X., Hu, L. Q., Yang, L. H., Zhang, T., Dai, R. B., Zhang, Y. J., ... & Lin, W. H*. (2023). Cell Reports, 42(7).
Spatiotemporal formation of the large vacuole regulated by the BIN2-VLG module is required for female gametophyte development in Arabidopsis. Hu, L. Q., Yu, S. X., Xu, W. Y., Zu, S. H., Jiang, Y. T., Shi, H. T., ... & Lin, W. H*. (2023). The Plant Cell, 35(4), 1241-1258.
Ovule initiation: the essential step controlling offspring number in Arabidopsis. Yu, S. X., Jiang, Y. T., & Lin, W. H*. (2022). Journal of Integrative Plant Biology, 64(8), 1469-1486.
PIN3 positively regulates the late initiation of ovule primordia in Arabidopsis thaliana. Hu, L. Q., Chang, J. H., Yu, S. X., Jiang, Y. T., Li, R. H., Zheng, J. X., ... & Lin, W. H*. (2022). PLoS Genetics, 18(3), e1010077.
Interaction of brassinosteroid and cytokinin promotes ovule initiation and increases seed number per silique in Arabidopsis. Zu, S. H., Jiang, Y. T., Chang, J. H., Zhang, Y. J., Xue, H. W., & Lin, W. H*. (2022). Journal of integrative plant biology, 64(3), 702-716.
Asynchrony of ovule primordia initiation in Arabidopsis. Yu, S. X., Zhou, L. W., Hu, L. Q., Jiang, Y. T., Zhang, Y. J., Feng, S. L., ... & Lin, W. H*. (2020). Development, 147(24), dev196618.
Two tonoplast proton pumps function in Arabidopsis embryo development. Jiang, Y. T., Tang, R. J., Zhang, Y. J., Xue, H. W., Ferjani, A., Luan, S., & Lin, W. H*. (2020). New Phytologist, 225(4), 1606-1617.


 PDF
PDF SUPP INOF
SUPP INOF SUPP INOF
SUPP INOF